| �� HOME >�q�X�^�~���@�v���u�����\�A�����M�[�l�H���ł́u���������v�ɂ��ā\ |
 |

�q�X�^�~���@�v���u����
�\�A�����M�[�l�H���ł́u���������v�ɂ��ā\
 ���䌚�v�i�����Ɛ���w�j
1.�q�X�^�~�������ۂ̔���
���̃^�C�g���́u�q�X�^�~���@�v���u�����v�͎��̑��_����̉��t�ł���ؖ����v�搶�������ꂽ�����̃^�C�g�������肵�����̂ł���B�ؖ��搶�́A1947�N�i���a22�N�j�ɕ��ߎs�ɐV�݂��ꂽ���s��w�_�w�����Y�w�Ȃ̏��㋳���ł���B���ł͊w��̖��̂ɂ��p�����Ă���u�ۑ��v�Ƃ������t�̑n�o�҂ł�����A�w�H�i���s�w�x�i1944�N�A�͏o���[�j�Ɏ����ŁA1949�N�ɂ́w�H�i�ۑ��w�x�i���q���X�j�����s����Ă���B
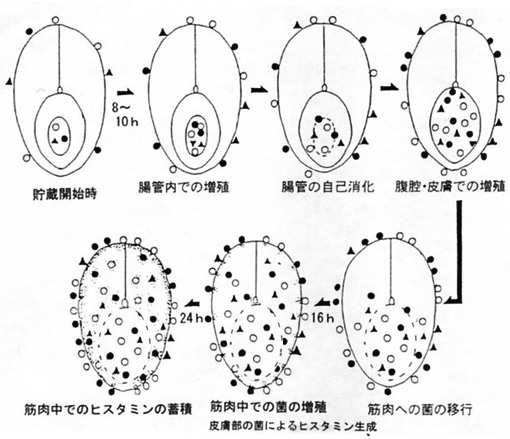
�@�搶�����߂ōŏ��ɍs��ꂽ�����́A�������Y���H�i����1�ʂ̐��Y�z�ŁA���s�A�H���łȂǂ̖��̑����������Y���萻�i�̕��s�ɂ��Ăł���A���ŃA�����M�[�l�H���łɊւ��Ăł������B
�@�A�����M�[�l�H���ł̌������n�߂�ꂽ�̂́A1947�N�̑n�݊Ԃ��Ȃ�����A����̋L�҃N���u�̐l�����������Č��w�ɗ���ꂽ�ۂ̒��H�̃T�o�ł��̐H���ł��N�������̂����������ł������炵���B�A�����M�[�l�H���ł��N�x�̒ቺ�����J�c�I�ȂǂŋN���邱�Ƃ͍]�ˎ��ォ��m���Ă������A�����Ԃ��́u���������v�͕s���ł������B�����e�n�Ŗ{���ł���������1950�N�㏉���ɂ́A���̐H���ł��q�X�^�~���ɂ��炵�����Ƃ͂��łɕ������Ă������A���̐��������̓����y�f�ɂ����̂��ۂɂ����̂��ɂ��Ă͕s���ł���A�����������̌����O���[�v�����̉𖾂������Ă������Ƃ��������\���ꂽ�_�����琄�@�����B
�@�ؖ����1951�N�����q�X�^�~�������Ɋւ��錤�����ʂ͓I�ɔ��\���Ă���A���̌�1960�N�܂łɖ�25��̊֘A�_���\���Ă���B�q�X�^�~�������ۂɊւ��錤���͂܂��A����a�Î���ɂ��1954�N�ȍ~�A�͒[�r������ɂ��1955�N�ȍ~�ɔ��\����Ă���A���ꂼ��7���15��̊֘A�_��������B
�@���̂����q�X�^�~�����������s�ۂɂ���Đ�������邱�Ƃ����߂Č��o�����͖̂ؖ��E�͍��i1953�N�j�ł���B�ؖ��炪�������������ۂ́A��ɖ{�H���ł̎�v�����ۂƂ��Ē��ڂ����Proteus morganii�i����Morganella morganii�j�ƍ����ł��������A�ቷ���B���Ȃǂ̓_�œ�����P. morganii�̋L�q�Ƃ͈�v���Ȃ���������Achromobacter histamineum�Ƃ��ĕ��ꂽ�B�H���œ��v�ł́A�A�����M�[�l�H���ł͌��݂����w���H���łƂ��ĕ��ނ���Ă��邪�A����͓����A���̐H���ł̌����������q�X�^�~���Ƃ������w�����ł��邱�Ƃ͔������Ă������A���̐��������ɂ��Ă͕s���ł��������Ƃ̖��c�ł��낤�B���̓_�ł��̃q�X�^�~�������ۂ̔����͈Ӌ`���傫���ƍl������B
�@�A�����M�[�l�H���łɊւ��Ėؖ��炪�s���������̗v�|�͎��̒ʂ�ł���B���Ō��������̎�̂͋����G�L�X���̃q�X�`�W����P. morganii�ɂ���ĒE�Y�_����Đ�����q�X�^�~���ł���B�]��Proteus �̎��K���x��35���t�߂ł���A�܂��q�X�`�W���E�Y�_��p�����邱�Ƃ��S������Ă��Ȃ��������ƂȂǂ���V��ƍl�������A���̌�ڍׂɌ����������ʁAP. morganii�Ɠ��肷��̂��ł��Ó��ł��邱�Ƃ𖾂炩�ɂ����B���̍ۂ͊C�Y���̐��������炷�łɋ��̕\�ʂ̔S�������ɏ�݂��i�S�ې���1/1000���x�j�A���̎��㑬�₩�ɑ��B���ăq�X�^�~����������B���̍ۂ͒����t�߂ōł����B���͑傫�����A�q�X�^�~���̐��Y�ʂ͏��Ȃ��A�_����ł͑��B�͊ɖ��ł��邪�A�q�X�^�~���̐��Y�ʂ͑����B��r�I�ቷ�ő��B�����ꍇ�ɂ̓q�X�^�~���̐��Y�ʂ͑����B�G�L�X���Ƀq�X�`�W�����قƂ�NJ܂܂Ȃ������̏ꍇ�ɂ́A���s���������x�ɂ܂Ői��łȂ���q�X�^�~���͐�������Ȃ��B�q�X�^�~�����ቷ�i6�`7���j�ł���������邱�Ƃ����o�����B���̂ق��A�ۂ̃q�X�`�W���E�Y�_�y�f�ɂĂ����炩�ɂ��A���̒���������ݒ肷�邤���ő���ȍv��������Ă���B�����̌����𒆐S�ɂ�����������Fish as Food, Vol.1�h(Academic Press, 1960)�ɏ�q�́gThe Histamine Problem�h�Ƃ����^�C�g���ł܂Ƃ߂��Ă���B
2.�C�m���q�X�^�~�������ۂ̔���
�q�X�^�~�������ۂɂ��Ă͊C�m���q�X�^�~�������ۂ̔������d�v�ł��낤�B����͎��̓������Y��w�ł̏�i�ł��������Ϗ����搶��ɂ����̂ł���B
�@�N���₻�̉��H�i�̃q�X�^�~�������ۂƂ��ẮA�]��M.morganii�CCitrobacter freundii, Enterobacter aerogenes, E. cloacae, Raoultella planticola�Ȃǂ̒����ۉȍۂ���ł���A���ۂ̐H���Ŏ��Ⴉ��̕�������M. morganii�ARaoultella planticola, Hafnia alvei�Ȃǂ̒����ۉȍۂł������B
�@�C�m�R���̃q�X�^�~�������ۂɂ��ẮA1981�N�ȍ~�A���ς�͋��̒ቷ�����Ɋւ���ڍׂȌ����ɂ���āA�C�m�⋛�̒��ǁA�̕\�Ȃǂɂ��ቷ���ƒ�������2��̍D�����q�X�^�~�������ہAPhotobacterium phosphoreum��P. damselae�����݂��邱�Ƃ𖾂炩�ɂ���Ă���B�O�҂͑��B���K���x��20���t�߂ɂ���A2.5���ő��B���邪35���ł͑��B�ł��Ȃ��ቷ�ۂł���B�܂��H�����Y���̔|�n�ł͑��B�ł����A���K�H���Z�x����2���̍D���ۂł���B�܂���҂�P. damselae��35���ő��B���邪4���ł͑��B�ł��Ȃ������ۂł���B
3.�C�m���q�X�^�~�������ۂ̏d�v��
�����ۉȂ̃q�X�^�~�������ہiMorganella morganii�Ȃǁj��2��̊C�m���q�X�^�~�������ۂ̉��݊C��ł̏o���ׂ�����(�}1)�ł́A�ቷ�D�����̃q�X�^�~�������ہiP. phosphoreum�j�͓~���珉�Ăɂ����đ��݂��A�Ăɂ͒����D�����̋�(P. damselae)�������o������B���M. morganii�͐���C���i���͘p�j����͌��o���ꂸ�A��r�I���ꂽ�C���i�����p�p�����j����ċG�Ɍ��o�����B���������ċ��l����̑N���Ɏ�ɕt�����Ă���͍̂D�����q�X�^�~�������ۂ̕��ł���AM. morganii�͋��l�㐅�g�����̐��₻�̌�̗��ʉߒ��ł̉�������ƍl������B
�@�܂��A�s�̑N���ɂ��Ē��ׂ���(�}2)�ł́A�����D�����ۂ�M. morganii�͎�ɉď�Ɍ��o����A�ቷ�D�����ۂ͎��N���p�x�Ɍ��o�����B���̂����A�����D�����̃q�X�^�~�������ۂ͉Ă̑N�����瑽���Ƃ��ɂ�103�`104/cm2���o����邱�Ƃ�����A������M. morganii�Ɠ����x�ɋ����q�X�^�~�������\�����̂ŁA�ߋ��̐H���Ŏ���̒��ɂ͖{�ۂɂ����̂��܂܂�Ă����\��������B����A�ቷ�D������P. phosphoreum��2.5�������̋������ɑ��ʁi61�`144mg/100g�j�̃q�X�^�~�����Y�����邱�Ƃ��m�F����Ă���̂ŁA�ቷ�������̑N���ɂ�����q�X�^�~�������ɂ͖{�ۂ��֗^���Ă���\���������B�]��P. phosphoreum�ɂ��H���ł̕�͂Ȃ��������A����͖{�ۂ������Ɏア���ƂƁA��ʂɗp�����Ă��錟�������i�H�����Y���A35���|�{�j�ł͌��o����Ȃ����߂Ǝv���A�@�����ΐH���Ŏ���ׂĂ݂����ƍl���Ă������A��w�ł͐H���ŃT���v���̓��肪��������ł��Ȃ������B���̌�A�{�ۂ�2004�N�ɐ_�g���j����ɂ��ۊ������킵�ɂ��H���Ŏ��Ⴉ�番������Ă���B
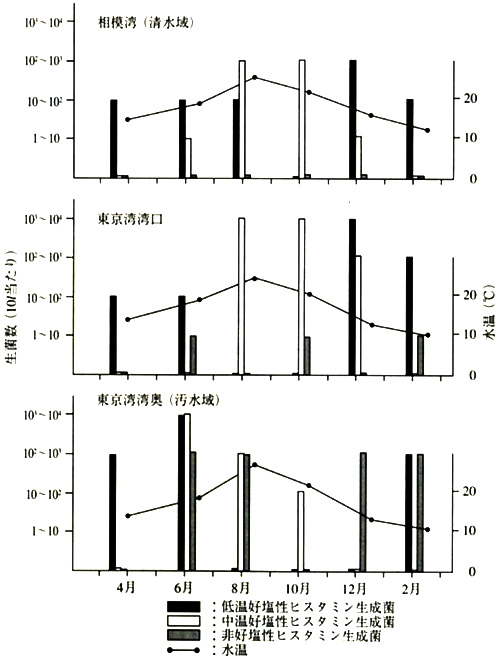
�}1�D�C�����̃q�X�^�~�������ې��̋G�ߕω��i�^����A1990�j
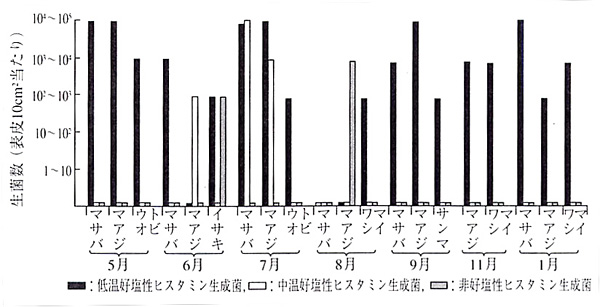
�}2�D�N���ɕt�����Ă���q�X�^�~�������ې��̋G�ߕω��i�^����A1990�j
4.�������ł̃q�X�^�~���~�ς̗l�q
P. phosphoreum�͋��̔畆�����łȂ��A���Ǔ��e���ɂ��ʔN���o����A���̋ې��͒������̐����ۂƓ�����10�`102�{�قǑ������݂���̂ŁA�ؓ����ł̃q�X�^�~���~�ςɂ͒��Ǔ��̐����ۂ��d�v�ł���B�}�T�o��25���Œ��������ۂ̃q�X�^�~�������ۂ̋����ׂ����ʁi�}3�j�ł́A���Ǔ��̐����ۂ�8�`10���ԂŒ��������B���A���̌㒰�ǂ̎��ȏ����ŕ��o���֊g�U���A����ɖ��ۂł������ؓ����ֈڍs�A���B���A�q�X�^�~�����E�~�ς���B���Ǔ��ɂ͑����Ƃ��ɂ�107�`108/g�̃q�X�^�~�������ۂ����݂���ꍇ������A���̂悤�ȋ��̂ʼn��x�Ǘ��̕s��ۂɂ���ē����̎��ȏ����������i�s�����ꍇ��A�����������s�\���ȏ�Ԃŕ��u�����悤�ȏꍇ�ɂ́A�������ɂ��Ȃ�̃q�X�^�~�����~�ς��A���ꂪ�����E�𓀂��J��Ԃ����Ƃł���ɋؓ����ֈڍs����\��������B���������ċ��l��ɓ������������邱�Ƃ́A���̌�̒������̃q�X�^�~��������h���_�ňӖ������낤�B
�}3�D�N���̒������i25���j�ɂ�����q�X�^�~���~�ς̋@�\�i�^����A1990�j
�}�͋��̒f�ʂŁA�~�͓������珇�ɒ��ǁA���o���A�\��������A
�}���̎��Ԃ͒����J�n������̌o�ߎ��ԁA���FP.phosphoreum,���FP.damselae,���F�����K����
5.�q�X�^�~�������̓����y�f���ɂ���
�������ł̃q�X�^�~���������A�q�X�^�~�������ۂł͂Ȃ��A�����̓����y�f�ɂ���Ă��邩������Ȃ��Ƃ��������ɂ�����ُ���iabnormal meat�j�̖�肪����B�ُ���Ƃ́A���Ƃ��āA�q�X�^�~����543mg/100g�ƈُ�ɒ~�ς��Ă���Ⓚ�L�n�_�}�O���ɂ����āA�q�X�^�~�������ۂ�10�`100/g�������݂����A�������ُ틛�����̃q�X�`�W���E�Y�_�y�f���������틛���ɔ�ׂėL�ӂɍ����i�q�X�^�~���������ؓ��R���y�f�ɂ��\��������j�Ƃ����悤�Ȏ���ł���B�������A���̂悤�Ȏ���͍D�����q�X�^�~�������ۂ������Ɏア���߁A�����T���v���ł͎��ł��Ă��܂��ƍl����Ɛ��������B
�@�}4�͍D�����q�X�^�~�������ۂɑ��铀���̉e���ׂ���ł��邪�AP. damselae�̋ۑ̌����t���|20���Œ��������ۂ̐��ې��ƃq�X�`�W���E�Y�_�y�f�����̕ω��ׂ����ʂł���B���̋یQ�͓����ɋɂ߂Ďキ�A�ې��͓���7����ɂ�1/108�ȉ��ɋ}�����邪�A�ۉt�̍y�f������7����ł���50�����ێ�����Ă���B�ُ���ɂ����鍂���y�f�����͂��̌��ʂ�������ł���̂ł͂Ȃ��낤���B
�}4�D�|20����������P. damselae�̐��ې��ia�j�ƃq�X�`�W���E�Y�_����(b)�̕ω��iFujii��A1994�j
�@�@
�Q�l����
�{�e�ŐG��Ȃ����������╶���͎��ٍ̐e���Q�Ƃ��������B�Ȃ��q�X�^�~���H���łɂ��ẮA�{�����}�K2009�N11�����ɂ��f�ڂ��Ă���̂ŕ����Ă������������B
(1)���䌚�v�F �q�X�^�~��������,�wHACCP�Ɛ��Y�H�i�x�i����E�R���ҁj,�@p.59-74, �P���Ќ����t (2000).
(2)���䌚�v�F�A�����M�[�l�H����, ���{�H�i�������w��G��, 23, 61-71 (2006).
(3)���䌚�v�F�ې��H���łƂ��ẴA�����M�[�l�H����, �H�i�q���w�G��, 47�CJ343-J348 (2006).
����
���s��w��w�@�_�w�����Ȕ��m�ے��C���A���Y�����C�搅�Y���������������������A�������Y��w�E�����C�m��w�����Ȃǂ��o��.�A���ݓ����Ɛ���w���C�����E�����Ȋw���������B�_�w���m.�B
��啪��F�H�i�������w�B
|

