最近、ガス置換包装、真空包装、脱酸素剤及びアルコール製剤使用包装が普及するに伴い、また防腐剤及びアルコールがよく用いられるようになってから従来問題とならなかった食品に酢酸エチルが生成し食品を変敗させる事例が多くなってきた。つまり、セメンダイン臭、シンナー臭の生成である。原因菌は殆どPichiaanomalaであり、一部Candida、Trulopsisが関与する。酢酸エチル生成により変敗した食品の共通点は食品保存料、風味改良剤としてエタノール及び酸を生成する。従って脱酸素された含糖食品はこれらの菌によって変敗しやすい。Pichiaanomala はエタノール資化性が強く、酢酸エチルを生成する。
最近、製造2〜3日後の饅頭に石油臭が生成する現象が多発している。饅頭の主原料が小麦粉、生あん、砂糖、水飴、卵、粉乳、マ−ガリン、ソルビト−ル、膨張剤、シナモン抽出物、シナモンが入った保存料である場合に多く石油臭が発生している。石油臭の原因は石油ではなく、スチレン物質による。シナモン抽出物の主成分であるケイ皮酸がSaccharomycescerevisia,Pichiacarsonii,Candidafamata等の酵母の脱炭酸酵素により脱炭酸されてスチレンが生成することによる。シナモン抽出物はクスノキ科のニッケイから抽出されたもので、いわゆる天然添加物の一つで、ケイ皮酸その他種々の化学成分を含有し特有の臭気があり、また抗菌性や抗酸化性があるのでよく保存料に用いられている。
洋菓子の中で酵母汚染を受けやすい製品は水分が20%以上のもので、5%以下のビスケット、クッキ−、クラッカ−等はほとんど心配がない。例外的に水分5〜7%のクッキ−にSaccharomycescapsularia,Pichiaanomalaに由来する黒色斑点が生成した例があるが、この原因は製造工場の空中浮遊酵母による二次汚染であった。これは缶に詰めたクッキ−の表層の水分が一時的に上昇して、部分的に水分活性が上昇し、上記酵母が生育して黒色斑点が生成したものと思われる。黒色斑点の生成したクッキ−に存在する微生物は大部分がSaccharomycescapsularia,Pichiaanomalaの酵母であり、圧倒的にSaccharomycescapsulariaが多いことから、クッキ−の黒色斑点は本菌に由来する。また本菌は製造工場の天井、壁及び空中浮遊菌から検出されたところから製造工程中における二次汚染である。この変敗現象が多発してから、今までほとんど二次汚染菌のことを考えていなかった多くの干菓子工場おいてクリ−ンにつとめるようになった。
現在、農産加工食品の酵母による異臭生成現象が多発している。最も多いのがシンナ−臭やセメダイン臭であり、主成分は酢酸エチルである。いなりずし、ちらしずし、福神漬、たくあん漬、らっきょ漬、ピザパイ、生パン粉,ジャムのシンナ−臭はPichia anomalaに起因する。これらの原因酵母はほとんどがエチルアルコ−ルを資化するためにシンナ−臭の原因である酢酸エチルを生成する。このためシンナ−臭の生成した食品を製造している工場のほとんどは工場殺菌剤として又は原材料としてエチルアルコ−ルを使用している。応急措置としてエチルアルコ−ルの使用を停止すればシンナ−臭の生成はおきない。
佃煮は、れんこん、ごぼう、豆類、ふき、さんしょ、とうがらし等の農産物及び魚、貝類、海草類等の水産物又はそれらの加工品を原料として、これに醤油、水飴、砂糖等の調味料を使用し、煮詰めて濃厚な調味を施し、夏期おいても、少なくとも20日間程度の保存性を付与した調理加工食品である。佃煮の貯蔵効果としては、浸透圧、脱水作用等の物理的な原理を応用したものである。塩分あるいは調味料の濃淡により製品の味や保存性が左右され、特に生鮮原料の場合は農産物、水産物を問わず水分が多いために酵母によって変敗を起こしやすく、またこれらの原料の鮮度の低下によりて製品の価値が失われやすい。佃煮を調味料で分類すると3つに分類される。一番目は醤油、食塩が主体で甘味料は微量しか使わないものであり、ふき佃煮、はとう芥子佃煮、昆布佃煮、海苔佃煮、貝類佃煮、はぜの佃煮、生アミの佃煮等がある。二番目は醤油、砂糖、水飴、食塩を併用しているもので、はぜやふなの甘露煮、かつおやまぐろの角煮、わかさぎの佃煮、こうなごの佃煮、削節佃煮、混合品佃煮がある。三番目は醤油を使わず砂糖、水飴を主体とし微量の食塩を加えて調味したもので、スルメ佃煮がある。これらの種類により生育する酵母は変化する。佃煮の酵母による変敗は昆布巻きのHansenula subpelliculosa,Pichia anomalaによる白斑点生成及び膨張、海苔佃煮のRhodotorulaによる淡橙色斑点生成、海苔佃煮のPichia anomala等による異臭、液化、れんこん佃煮のTorulopsisによる白色小斑点、キンピラ佃煮のSaccharomycesによる異臭、緑変、昆布佃煮のPichia anomalaによる酢酸エチル臭、タケノコワカメ佃煮のTorulopsisによる白色小斑点、葉とうがらし佃煮のCandidaによる白色斑点等極めて多い。今までは酵母による佃煮類の変敗は比較的少なかった、しかし低塩、低糖化が進み、脱酸素剤、粉末エタノ−ル製剤等の使用が多くなったために酵母が主原因となる変敗が多発してきた。これは包装形態の変化によりいままで問題とならなかった酵母が生育して微生物菌叢が変化したためである。昆布巻の酵母による白斑点生成及び膨張現象について説明する。昆布佃煮の原料とされるのは真昆布、利尻昆布、細目昆布、長昆布等であるが、とりわけ長昆布と利尻昆布が多く利用されている。この原藻である昆布には5.7×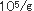 の細菌と1.2× の細菌と1.2×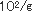 の酵母が付着している。水洗あるいは酢酸処理により菌数は若干減少するが、次の昆布巻工程や包装工程において二次汚染を受けて増加する。次の加熱工程により菌数は減少するが、加熱工程以前の菌数により残存菌数は決定され、pHが5.0〜5.2、水分55〜60%、水分活性0.92〜0.95であるため通常著しい変化はみられない。ただ物理的現象として、親水性多糖類が構成成分であるため周囲の湿度により吸脱湿して、外観が損なわれることがある。従来、昆布佃煮、スルメ佃煮、混合佃煮、でんぶといった濃口佃煮は、概して、糸状菌により変敗する場合が多く、Aspergillus glaucus,Aspergillus niger,Penicillium,Rhizopus,Mucorが主汚染菌であった。最近の包装形態等の変化により濃口佃煮でも、吸湿が著しい場合には酵母の発育がみられ、また浅炊き佃煮の変敗は主として酵母の発育による場合が多い。白斑点が生成して膨張した昆布巻の場合、Bacillus cereus,B.subtilis,B.macerans等の細菌が5.0× の酵母が付着している。水洗あるいは酢酸処理により菌数は若干減少するが、次の昆布巻工程や包装工程において二次汚染を受けて増加する。次の加熱工程により菌数は減少するが、加熱工程以前の菌数により残存菌数は決定され、pHが5.0〜5.2、水分55〜60%、水分活性0.92〜0.95であるため通常著しい変化はみられない。ただ物理的現象として、親水性多糖類が構成成分であるため周囲の湿度により吸脱湿して、外観が損なわれることがある。従来、昆布佃煮、スルメ佃煮、混合佃煮、でんぶといった濃口佃煮は、概して、糸状菌により変敗する場合が多く、Aspergillus glaucus,Aspergillus niger,Penicillium,Rhizopus,Mucorが主汚染菌であった。最近の包装形態等の変化により濃口佃煮でも、吸湿が著しい場合には酵母の発育がみられ、また浅炊き佃煮の変敗は主として酵母の発育による場合が多い。白斑点が生成して膨張した昆布巻の場合、Bacillus cereus,B.subtilis,B.macerans等の細菌が5.0×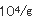 であり、主原因とは考えにくく、Pichia anomala,Hansenula subpericulosa等の酵母が2.6× であり、主原因とは考えにくく、Pichia anomala,Hansenula subpericulosa等の酵母が2.6×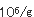 検出されたところから主原因菌と考えられた。このように変敗品は原料の上記酵母の汚染が著しく、工場からの上記酵母の二次汚染が多いことと蒸気殺菌が不十分であったためであると考えられた。最近、瓶に詰めた海苔佃煮に異臭が生成し、液化するとともに味が変化する現象が多発し、その原因を検討した結果工場からの二次汚染に由来する乳酸菌Weisella viridescenceが原因菌であった。同時に一部の製品に淡橙色斑点が生成するという現象があったので検討すると、Rhodotorula属の酵母であった。このRhodotorulaは調味料混合工程、蒸熟工程、瓶詰工程より検出されたところから工場からの二次汚染が原因であった。海苔佃煮工場は原藻の洗浄に多くの水を使用するために湿度が高く、さらに原料が多種にわたるために微生物汚染は著しい。海苔佃煮の原料、特に糖類や調味料を多量に使用するため、またこれらが工場に多く散乱するために床や機械等に付着した汚染菌が多い。工場では長年の間、床等の殺菌に300ppmの次亜塩素酸ナトリウム液を散布してきたが大腸菌群、乳酸菌、酵母の増殖が認められたところから、これらの菌が長年の同一の殺菌剤に使用により耐性菌が出現したと考えられた。 検出されたところから主原因菌と考えられた。このように変敗品は原料の上記酵母の汚染が著しく、工場からの上記酵母の二次汚染が多いことと蒸気殺菌が不十分であったためであると考えられた。最近、瓶に詰めた海苔佃煮に異臭が生成し、液化するとともに味が変化する現象が多発し、その原因を検討した結果工場からの二次汚染に由来する乳酸菌Weisella viridescenceが原因菌であった。同時に一部の製品に淡橙色斑点が生成するという現象があったので検討すると、Rhodotorula属の酵母であった。このRhodotorulaは調味料混合工程、蒸熟工程、瓶詰工程より検出されたところから工場からの二次汚染が原因であった。海苔佃煮工場は原藻の洗浄に多くの水を使用するために湿度が高く、さらに原料が多種にわたるために微生物汚染は著しい。海苔佃煮の原料、特に糖類や調味料を多量に使用するため、またこれらが工場に多く散乱するために床や機械等に付着した汚染菌が多い。工場では長年の間、床等の殺菌に300ppmの次亜塩素酸ナトリウム液を散布してきたが大腸菌群、乳酸菌、酵母の増殖が認められたところから、これらの菌が長年の同一の殺菌剤に使用により耐性菌が出現したと考えられた。
水産加工食品はかまぼこ、ちくわ等の練製品、佃煮、塩から等の塩蔵品、するめ等の乾製品、かつお節等のくん製品、水産缶詰、海草加工品と多岐にわたっている。
水産加工品においても塩から等の塩蔵品等は酵母による香気付与等に重要な役割を果たしている場合もあるが、酵母により異臭、斑点生成、容器の膨張等の有害な現象を引き起こしている。
イカ塩辛、ウニ塩辛、カツオ塩辛、イカ燻製、タコ燻製、魚肉ソ−セ−ジ等の水産加工食品の変敗品よりTorulopsis,Rhodotorula,Zygosaccharomyces,Debaryomyces等の酵母が多く検出されている。その原因菌は亜硝酸塩を資化できるDebaryomyces hannseniiが多いのが特徴であるが、Candida,Trulopsis,Trichoderma等も多く報告された。イカ燻製の白斑点生成はCandida lactis-condensiに起因し、海苔巻きすしにみられる白斑点は具及びすだれ、手袋に由来するSaccharomycesであった。かまぼこ等の練製品の変敗は主として細菌であるが酵母も同時に生育して変敗原因菌の一つになりうる。かまぼこからTorula nukaiso,Torula ohtaniが分離され、変敗したかまぼこの表面の微生物菌叢の2%が酵母であり、ネトの部分に多数の酵母が生育し、それらの多くはCandidaであった。この原因菌は海水からRhodotorula, Torulopsis, Cryptococcus, Saccharomyces, Sporoboromyces及びCandida等の多くの酵母が検出されているところから原料魚からの一次汚染及び製造工場からの二次汚染の両者が考えられる。イカ塩辛、ウニ塩辛、カツオ塩辛等の水産発酵食品よりTorulopsis,Rhodotorula,Zygosaccharomyces,Debaryomycesが分離されている。ソ−セ−ジ類には殺菌が不十分であると酵母が生育してスライムの生成原因菌となることが知られている。その原因菌は亜硝酸塩を資化できるDebaryomyces hanseniiが多いのが特徴であるが、Candida,Torulopsis等も報告されている。水産食品においてもシンナ−臭が生成して大きな問題となっている。竹輪、はんぺん、カニかまぼこ、かまぼこのシンナ−臭はいずれも工場の製造工程より検出されたPichia anomalaに起因する。本菌はエタノール資化性が強く、酢酸エチルを生成する。その他アルコールを資化して酢酸エチルを生成する酵母はTrulopsisi,Candidaがあり、これらの酵母によりシンナー臭が出現しているケースもある。
香辛料に由来するスチレンによる石油臭はいかの燻製、竹輪、カニかまぼこ、はんぺん、肉まんにおいて生成し、原因菌はPichia carsonii,Candida famata,Debaryomyceshansenii,Torulopsisの4種類が圧倒的に多い。石油臭の原因は石油ではなく、スチレン物質による。最も多いのはシナモン抽出物の主成分であるケイ皮酸が酵母の脱炭酸酵素により脱炭酸されてスチレンが生成することによる。
「香り」は食品の品質を決定する重要な因子で、味と同様に食品の評価に大きく影響する。発酵食品においては香りは最も重要な因子であり、その製造では異臭等のオフフレーバーの生成を抑制し、好ましい香りを生成させるように研究開発されてきた。しかし異臭の生成による発酵食品の変敗は多い。発酵食品にバナナ臭がするのは酵母が生産する酢酸イソアミルであり、酢酸イソアミルはイソアミルアルコールが前駆体となり、アルコールアセチルトランスフェラーゼの働きでアセチル-CoAからアセチル基が転移することにより生成する。酢酸イソアミルの前駆体であるイソアミルアルコールを酵母に添加すると、細胞が異常に伸長して、細胞分裂が遅延する。イソアミルアルコール高生産株でも同様の形質の変化が認められるところから、細胞内のイソアミルアルコールの濃度上昇は酵母にとって好ましくない。このため細胞内のイソアミルアルコール濃度が高まると、これを細胞外に排泄して毒性を低減するか或いはアルコールアセチルトランスフェラーゼによって酢酸イソアミルに変換してイソアミルアルコールの毒性を低減している。リンゴ様の香りは「カプロン酸エチル」であり、同様の酵素機作で生成する。異臭生成は酵母にとって細胞増殖をスムーズに進めるための解毒作用という重要な役割を持っている。
発酵臭といえば、酢酸イソアミルやカプロン酸エチル等が中心であるが炊飯米が傷んだ時の臭いジアセチル臭がオフフレーバーとして生成する。発酵食品ではピルビン酸からエチルアルコールが生じると同時に、分岐鎖アミノ酸を生合性するため、ピルビン酸はアセト乳酸シンターゼによってα―アセト乳酸に変換される。α―アセト乳酸は非酵素的に酸化されてジアセチルとなり、アルコールデヒドロゲナーゼによりアセトインを経て無臭の2,3ブタンジオールに代謝されるが、これらの代謝副産物は極性が低く、細胞膜を拡散により透過できるため細胞外にも存在する。発酵中は酸素分圧が低いためにジアセチルの生成は抑制される。しかし酸素分圧が高まる食品の上部に酵母が存在するとジアセチルの生成が促進される。このため、α―アセト乳酸濃度が高くなるとジアセチル臭が生成する。ビール醸造においてもアセト乳酸シンターゼを阻害することによりα―アセト乳酸の生成を抑制してジアセチルの生成を抑制しているこれらの技術を採用することにより。今までジアセチル臭が強いために異臭生成品として処理されてきた発酵食品が減少すると考えられる。
ビールは、数千年の長い歴史を経て、一般に抗菌性のある安全な飲料へと進化してきた。この原因はアルコール(4〜5.5%)を有し、低pH(pH4.0〜4.5)で溶存酸素濃度(0.1〜0.3ppm)が低いこと、栄養源(発酵性糖含量0.5%以下)が少ないこと及びホップ中に含まれる抗菌成分(グラム陽性菌に抗菌力)を含有することによる。病原性細菌はビール中では生育することができず、死滅していく。
淡色ビールに接種した病原性細菌、大腸菌は7日後、サルモネラは3日後、黄色ブドウ球菌は3日後には完全死滅する。自然界に存在する微生物がビール中で生育し、ビール本来の香味や清澄度に影響を与える。これらの微生物をビール有害菌といい、ビール醸造工程では特にこれらの有害菌を中心とする微生物管理は重要である。ビール有害菌は次の3つの微生物群に分類される。1つ目は乳酸菌(Lactobacillus brevis, Lactobacillus limdneri, Pediococcus damnosus),2つ目は偏性嫌気性細菌(Pectinatus frisingensis, Pectinatus cerevissiphilus, Megasphera cerevisiae), 3つ目は酵母(Saccharomyces diastaticus、Saccharomyces ellipsoideus, Saccharomyces byanus, Saccharomyces willianus,Saccharomyces logos、これらの酵母は現在いずれもSaccharomyce cerevisiaseに分類されている。)である。乳酸菌は味を劣化させ、偏性嫌気性菌は混濁させ、酵母は風味を劣化させる。特にビール本来の香味を維持できる非加熱処 理のビール、いわゆる生ビールの微生物管理は重要であり上記3種類の微生物により変敗することがある。最近では市販の瓶ビールで沈澱物が異物として検出され、その原因が酵母であった。加熱ビールにおいても異物として酵母が検出される場合がある。発酵性糖含量が0.5%以上のビール中ではBrettanomyces,Candidaが増殖して混濁、沈澱等の原因となる場合がある。
醤油は大豆と麦類を主原料として麹を造り、塩水中で発酵熟成させた液体調味料である。発酵中、原料に含まれるタンパク質やでんぷんは麹菌の生産した酵素により低分子化される。また耐塩性酵母等により多様な成分が造られる。醤油は高濃度の塩分、低Aw及び各種成分により高い抗菌性を有している。しかしZygosaccharomycesやTorulopsisの耐塩性酵母により醤油表面に膜を形成して生育する。これらの酵母は醸造にかかわる酵母と同種で、不快臭や有害物質の生産はないが、外観等によりクレーム製品となる。
醤油は開栓後、常温で保存される場合が多く、開栓後もある程度酵母の生育を抑制できる品質であることが必要である。通常、醤油は80℃以上の加熱工程を経て製品化される。殺菌の他に酵素失活及び火入れ香の付与が目的である。ここで酵母はいったん完全に殺菌される。火入れ後、容器に充填し製品となる。充填工程には開放部があるために酵母が二次汚染して製品が劣化する。このために充填直前に火入れをするか、あるいは充填工程の無菌化を促進することが必要となる。通常、酵母の殺菌には50〜60℃で10〜15分間、70℃で2〜3分間、80℃で2〜3秒処理が必要である。
希釈用つゆで3倍希釈以上(塩分9%以上)の製品の場合、塩分が高いために酵母と乳酸菌による変敗が問題となる。2倍希釈以下のつゆは成分値調整により酵母生育を防止することは難しい。この場合は、加熱殺菌と、高温の状態(75〜85℃)で容器に充填する必要がある。充填後、容器を反対にしてキャップ部も加熱殺菌する。
高品質の醤油を製造することを目的として、醤油酵母Zygosaccharomycesrouxiiの培養液をスターターとして諸味に添加し、発酵を調整することが検討されている。多くの研究者による諸味への大量の醤油酵母菌体の添加による懸濁状態でのアルコール発酵でのpHは4.9〜5.4であり、酸素濃度、醸造期間により酵母菌数の著しく異なることが知られている。 |

